Identity and targeting
The first layer asks what the biological object is, how it is named, where it belongs, what it maps to, and which target, pathway, cell, system, or disease context it touches.
QTOS undercarriage
BioAtlas Intelligence is the visible platform. QTOS is the deeper physics-first framework beneath it — a proposed architecture for modelling biological state as layered coordination drift across timing, signalling, identity stability, adaptive flexibility, and systems-level attractor behaviour.
QTOS is presented here as a proposed systems-state modelling framework, not as a validated clinical diagnostic, treatment protocol, or established biological physics law.
Why physics first
Most biomedical systems begin with genes, pathways, symptoms, or biomarkers. QTOS begins from systems-state features: timing, charge, rhythm, geometry, coherence, energy flow, and state transition.
The public page introduces the architecture without releasing the protected manuscript logic, intervention matrices, raw diagrams, or diligence-only material.
Core thesis
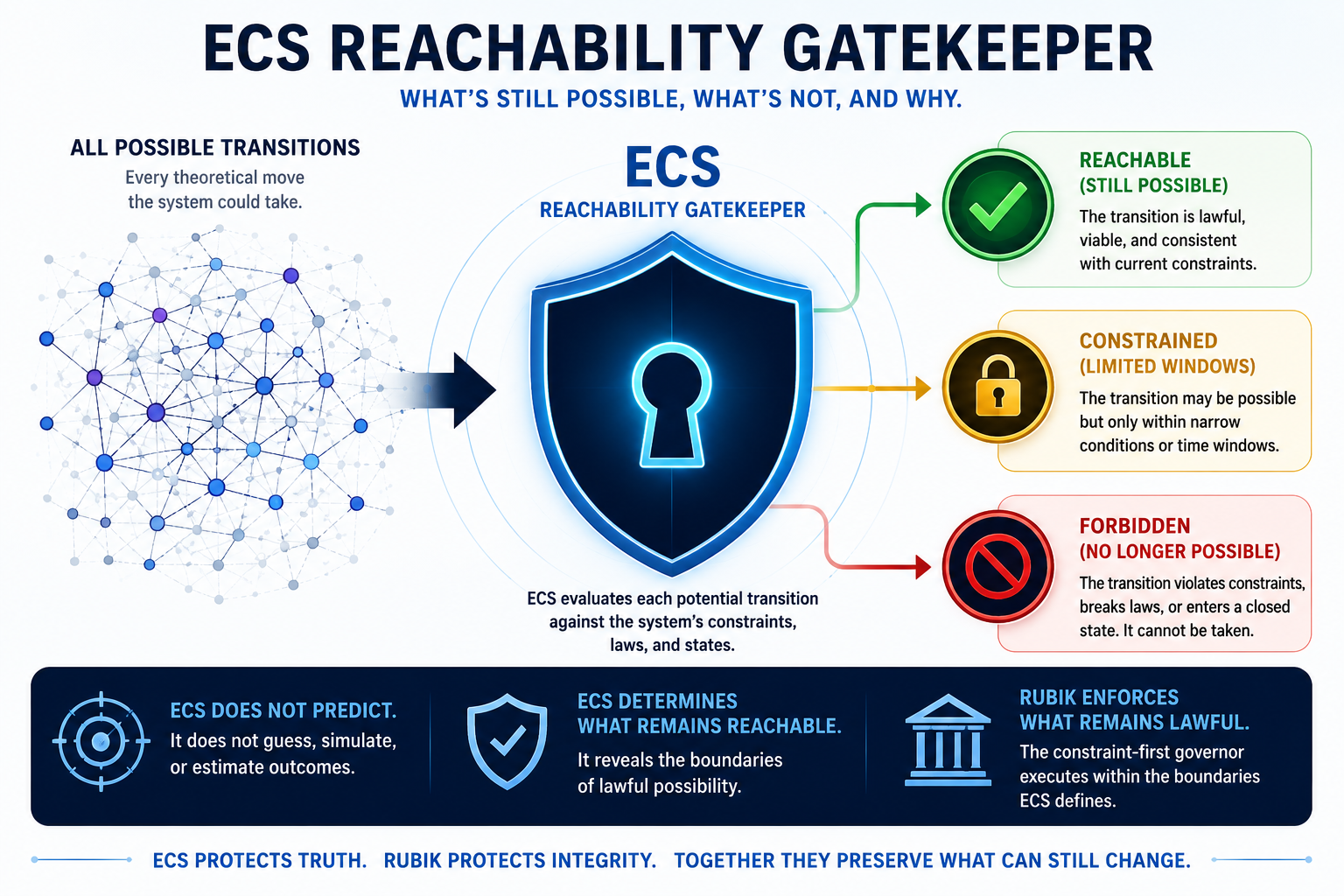
In a physics-first model, the decisive question is not only what molecules are present. It is what transitions remain reachable. Can the organism terminate threat, re-enter repair, restore timing, maintain energetic viability, and return to an integrated state without collapse?
ECS gatekeeper layer
QTOS places the endocannabinoid system near the centre of the model because ECS signalling participates in stress-axis termination, network excitability, inflammatory-state control, and energetic viability. In reachability terms, ECS integrity helps determine whether recovery windows remain open, narrow, or close.
Coherence-State Model
QTOS approaches biology as a dynamic systems-state architecture rather than a collection of isolated symptoms, organs, or molecular events. Instead of asking only what has failed inside the organism, the framework asks how biological coherence changes across timing, signalling, identity stability, adaptive flexibility, and system-wide coordination over time.
The model represents the organism as a set of interacting subsystem coherence vectors whose combined states influence whether biology remains inside a stable recovery basin or transitions toward pathological attractor behaviour. Within this framework, disease progression is interpreted as a threshold-gated collapse process emerging from accumulated coherence drift across multiple interconnected systems rather than a single isolated failure point.
The QTOS Coherence-State Model is presented as a proposed systems-biology and theoretical-state representation intended for research, computational reasoning, and conceptual modelling. It is not a validated diagnostic equation, clinical scoring system, or patient prediction engine.
Core state vector
S(t) = { R(t), I(t), C(t), F(t), P(t), A(t) }Rhythmic Timing Coherence
Immune / Inflammatory Coordination
Cellular Identity Stability
Adaptive Flexibility
Predictive / Perceptual Coherence
Attractor Basin Resilience
Formal state-space definition
R,I,C,F,P,A ∈ [0,1] S(t) = [R(t), I(t), C(t), F(t), P(t), A(t)] S(t) ∈ [0,1]^6 ΔS(t) = ||S(t+1) - S(t)||₂
This notation treats each QTOS coordinate as a normalised research-model dimension. Small ΔS values describe relative state stability; larger ΔS values describe stronger modelled transition or perturbation between adjacent states.
Operational variable definitions
Each QTOS state coordinate is presented as a research-model dimension with candidate proxy domains. These are not fixed clinical inputs, diagnostic thresholds, or validated patient-scoring variables.
Temporal coherence
Circadian stability, timing regularity, HRV-derived rhythm measures
Immune coordination
Inflammatory marker balance, immune signalling coordination
Cellular identity stability
Differentiation markers, epigenetic stability indicators
Adaptive flexibility
Recovery dynamics, adaptive response measures
Predictive / perceptual coherence
Behavioural consistency, cognitive-state coordination
Attractor resilience
Resistance to undesirable state transitions
Subsystem vector
X_j(t) = [R_j(t), I_j(t), C_j(t), F_j(t), P_j(t), A_j(t)]
j ∈ { brainstem, PFC, limbic, gut, liver, immune, heart, microbiome, ECS }Each subsystem is represented as a local coherence vector. The listed subsystems are public-facing examples, not a closed clinical inventory.
Systems-state interpretation
QTOS interprets the organism as a continuously evolving state-space system whose stability depends on coordinated coherence across multiple interacting biological domains. Within this architecture, health is not treated as a binary condition, but as the ability of the organism to remain dynamically reachable, adaptable, and recoverable across changing internal and external constraints.
Collapse emerges when subsystem instability accumulates faster than recovery coherence can be restored, progressively narrowing biological reachability until pathological attractors become dominant.
Transition ladder
Dysregulation may emerge before visible tissue pathology.
System instability may precede diagnosis or biomarker clarity.
Health is adaptive recoverability across changing constraints.
Chronic states can stabilise into self-reinforcing patterns.
G(t) = Σ_j w_j X_j(t) where w_j ≥ 0 and Σ_j w_j = 1
Global coherence is modelled as the weighted contribution of subsystem states. The weights are model parameters, not fixed clinical constants.
D_k(t) = 1 if G_k(t) > θ_k for Δt ≥ τ_k D_k(t) = 0 otherwise
A gate opens only when a coherence dimension remains above threshold for long enough. Thresholds and dwell-times are theoretical calibration variables.
Q(t+1) = T(Q(t), G(t), D(t), Ω) T : State × Context → State Ω ∈ C, where C = external constraint set
The next QTOS layer/state depends on the current state, global coherence vector, active gates, and biological constraint field.
Pstable(t) ↔ coherent recovery basin Pcollapse(t) ↔ pathological attractor basin
These are model-output concepts for attractor-state reasoning, not patient-level probability claims.
Model pipeline
How this maps to the Five Layers
The Five-Layer Collapse Story can be read as the macroscopic biological expression of deeper coherence-state transitions occurring beneath the surface of the organism.
Minor rhythmic instability, adaptive strain, signalling variance, and early threshold stress.
Protective regulation weakens as subsystem vectors begin failing persistence thresholds.
Cross-system coordination degrades across immune, metabolic, neurological, and signalling domains.
Collapse states stabilise into persistent self-reinforcing biological patterns resistant to recovery.
The pathological state develops persistence dynamics that may become increasingly self-stabilising over time.
Six-Layer Biophysics Intelligence is the shared interpretation architecture underneath BioAtlas. It is the public-safe form of the older databar/field architecture: a way to read biological objects through identity, mechanism, signalling context, regulatory memory, review relevance, and disease-state navigation instead of treating them as flat records.
The first layer asks what the biological object is, how it is named, where it belongs, what it maps to, and which target, pathway, cell, system, or disease context it touches.
The second layer asks how the object behaves: what switches it, what thresholds matter, what activates or suppresses it, and how it changes under stress, timing, exposure, or state pressure.
The third layer reads the surrounding signalling environment: receptor tone, ECS/GPCR/ligand context, immune signals, metabolic pressure, cytokines, redox state, and cross-system communication.
The fourth layer connects the object to epigenetic control, miRNA context, chromatin state, identity memory, timing drift, repair history, and prior biological state.
The fifth layer explains why the object may matter for research, therapeutic review, protocol context, formulation logic, commercial evaluation, or professional interpretation without giving public treatment direction.
The sixth layer places the object into disease-state context: biomarkers, cell states, oncology overlays, PCD logic, metabolic adaptation, subtype pressure, tissue behaviour, and reviewed access pathways.
The same six-layer logic now appears across metabolic intelligence, PCD intelligence, enzyme intelligence, cell intelligence, hallmark intelligence, pathway intelligence, and protected graph workbenches. Public pages explain the architecture; protected systems hold the deeper tensors, workbooks, graph exports, protected reasoning, and governed review material.
BioAtlas semantic topology
QTOS provides the theoretical language for coherence, timing, reachability, collapse pressure, adaptive flexibility, and attractor behaviour. It attempts to describe how biological systems drift between stable and unstable states under pressure rather than viewing disease as isolated disconnected events.
BioAtlas takes those systems-state ideas and translates them into a computational topology framework. Instead of treating enzymes, pathways, inflammatory systems, metabolic systems, and repair systems as separate silos, BioAtlas represents them as interconnected semantic structures inside a graph-backed biological state space.
In practical terms, this means enzymes, diseases, compounds, and biological states can be represented as comparable topology vectors. Similarity relationships, state transitions, drift behaviour, and cross-system coupling can then be explored computationally rather than purely through static pathway diagrams.
The important architectural boundary is that QTOS and BioAtlas are not the same system. QTOS acts as a theoretical systems-state framework, while BioAtlas acts as the computational biology and semantic topology layer. The mapping graph only describes where relationships may exist between the two.
Topology mapping equation
QTOS axis: S(t) = { R, I, C, F, P, A }
BioAtlas axis: B(x) = { boundary, timing, identity, immune, metabolic, resilience }
Mapping graph: M : B(x) → S(t)S(t) represents the QTOS systems-state model. It describes the overall condition of a biological system through coherence, flexibility, identity stability, inflammatory coordination, perception, and resilience.
B(x) represents the BioAtlas biological topology vector. Instead of describing abstract systems-state behaviour, it describes measurable biological topology patterns across six biological axes.
The mapping function M : B(x) → S(t)does not claim that the systems are identical. It simply defines a relationship graph showing where biological topology behaviour may relate to QTOS systems-state behaviour.
Biological state vectors
BioAtlas uses a computational biological vector that is intentionally separate from the QTOS physics-state vector. The goal is not to force biology into physics equations, but to create a structured semantic representation that allows biological systems to be compared consistently.
Each biological object receives a six-axis topology profile spanning boundary integrity, timing and oscillation behaviour, identity stability, immune coordination, metabolic pressure, and resilience capacity. Together these form a biological “shape” that can be analysed computationally.
Once represented as vectors, enzymes, compounds, diseases, and biological states can be positioned inside a semantic state space. This allows BioAtlas to explore similarity neighbourhoods, cross-system coupling, topology gravity wells, recursive drift behaviour, and emergent biological patterns.
Rather than replacing conventional biology, this acts as an additional systems layer designed to help visualise relationships that are normally fragmented across immunology, metabolism, epigenetics, redox biology, repair systems, and signalling networks.
BioAtlas biological topology vector
B(x) = { B₁, B₂, B₃, B₄, B₅, B₆ }
B₁ = boundary
B₂ = timing
B₃ = identity
B₄ = immune
B₅ = metabolic
B₆ = resilienceSimilarity(x,y) = cosine_similarity(B(x), B(y)) Drift: B(t+1) = B(t) + ΔB_pressure(t) Distance: d(x,y) = ||B(x) - B(y)||₂ Stability: ΔB(t) = ||B(t+1) - B(t)||₂ Attractor: limₜ→∞ B(t) = Aᵢ when repeated drift converges
Similarity(x,y)compares how close two biological topology shapes are to one another. If two diseases, enzymes, or compounds have very similar vector shapes, BioAtlas treats them as neighbours inside the semantic state space. Distance and stability are geometry helpers for comparing state separation and short-term movement; they are not clinical diagnostic scores.
B(t+1) = B(t) + ΔB_pressure(t)describes topology drift over time. Biological systems are not static — inflammation, metabolic overload, redox stress, repair exhaustion, and signalling disruption can all gradually shift the system into a different biological state.
B(t) → Aᵢdescribes attractor convergence. If the same pressures continue repeatedly, the system may begin stabilising around a recurring pathological or adaptive pattern.
QTOS ↔ BioAtlas bridge
QTOS physics layers and BioAtlas biological topology layers remain separate source systems. The bridge only says where concepts relate.
Bridge function
M(Bᵢ) = Sⱼ where: Bᵢ = BioAtlas biological topology axis Sⱼ = QTOS coherence-state axis
The bridge function says that a biological topology axis may relate to a QTOS systems-state axis. For example, the BioAtlas timing axis may relate to QTOS temporal coherence behaviour.
This does not mean the two systems are mathematically equivalent. The bridge only acts as a translation layer allowing biological topology behaviour to be interpreted through the broader systems-state framework.
Architecturally, this separation is important because it keeps the theoretical QTOS layer independent from the computational BioAtlas topology engine.
timing
Maps by relationship to temporal coherence.
identity
Maps by relationship to cellular identity stability.
immune
Maps by relationship to immune / inflammatory coordination.
resilience
Maps by relationship to attractor basin resilience.
boundary
Maps by relationship to predictive / perceptual coherence.
metabolic
Maps by relationship to adaptive flexibility constraints.
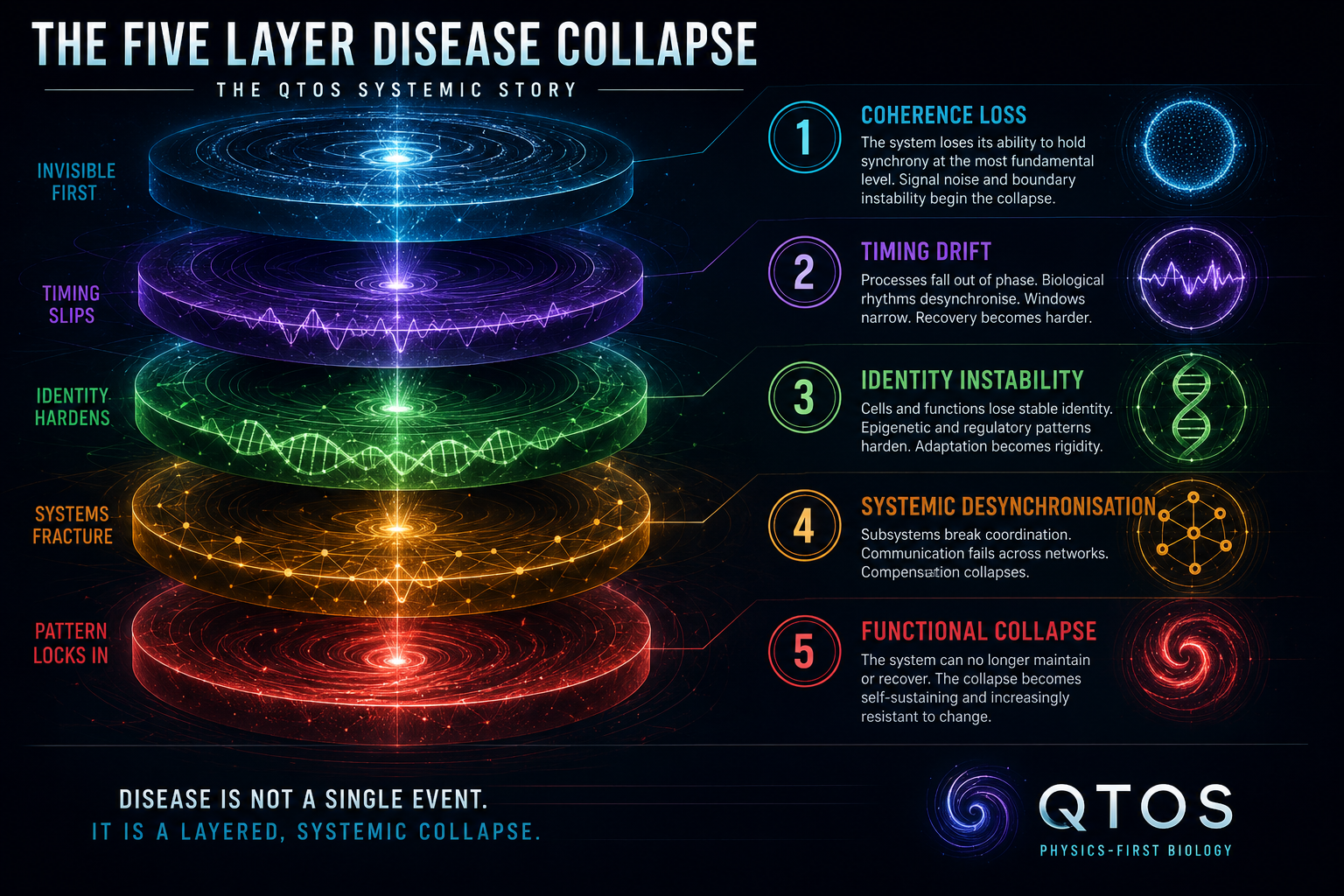
Five-layer model
The body does not suddenly become ill. It slowly loses order. First the boundary wobbles. Then time slips. Then identity hardens. Then systems stop talking to each other. And in advanced states, the collapse may become self-sustaining.
The five layers below are a public-safe preview of that story: no chapter numbering, no clinical instructions, and no protected book logic exposed.
Systems Layer 1
Coherence Boundary · ⊙ Guardian
Simple metaphor
Static appearing on a radio before the broadcast cuts out.
Before symptoms, before biomarkers, before visible damage, the body's organising signal begins to lose clarity. The system still works, but the quality of the internal message begins to degrade.
Imagine the body as a beautifully tuned orchestra inside a protective sphere. Every signal has timing. Every cell knows when to speak, when to listen, and when to stay quiet. Layer 1 is the moment that protective order starts to wobble. Nothing may look wrong yet, but beneath the surface the fine timing of the system begins to blur.
Boundary stability, timing precision, signalling coordination, redox balance, structural organisation, ECS regulation, and early systems-state stability.
Often invisible at first. Later, people may describe subtle instability, poor resilience, fatigue, stress sensitivity, or a sense that something is 'off' without clear test results.
Layer 1 is the silent beginning. It explains why a system can look normal while its deeper signal quality is already degrading.
Systems Layer 2
Biological Time · ∞ Eternal
Simple metaphor
Traffic lights across a city falling out of sync. The roads still exist, but flow turns into congestion.
The body is not just made of chemicals. It is made of rhythms. Layer 2 is what happens when those rhythms stop lining up.
If Layer 1 is signal noise, Layer 2 is timing failure. The body may still have the right molecules, but they arrive at the wrong time. Energy is made at the wrong moment. Repair signals arrive late. Stress chemistry lingers too long. The system still has the parts, but loses the timing that makes the parts work together.
Redox rhythm, ATP pulses, mitochondrial pacing, enzyme-cofactor timing, metabolite clearance, circadian-metabolic coupling, calcium waves, and ECS timing.
Cyclic fatigue, wired-but-tired feelings, non-restorative sleep, irregular energy, stress sensitivity, brain fog, and fluctuating inflammatory or autonomic patterns.
Layer 2 makes the reachability idea understandable: health is not only what chemistry is present, but whether the organism can still transition back into repair and balance.
Systems Layer 3
Biological Identity · Ψ Psyche
Simple metaphor
A building's control system saving a corrupted emergency setting as the new default.
Every cell and system needs an identity. Layer 3 is where timing drift begins to disturb the body's ability to maintain the right state.
Every cell has a role. It knows whether it is repairing, dividing, resting, defending, communicating, or standing down. That identity is not only written in DNA. It is maintained by timing, chromatin rhythm, epigenetic flexibility, miRNA regulation, and the ability to switch states appropriately. When timing stays distorted for long enough, the body tries to create stability, but it may create rigidity instead.
Epigenetic flexibility, chromatin rhythm, miRNA timing, programmed cell death availability, state memory, differentiation signals, and cell-state plasticity.
Reduced adaptability, persistent patterns, harder recovery, chronic inflammatory or metabolic tendencies, and a sense that the body has become stuck in a mode it cannot easily leave.
Layer 3 is where temporary dysfunction can begin hardening into persistent biological identity. The system may stabilise the wrong pattern.
Systems Layer 4
System-Wide Coherence · ✧ Luminary
Simple metaphor
A city where the power grid, transport system, emergency services, internet, and water network are all running, but no longer coordinating.
Layer 4 is the point where hidden collapse becomes visible because the body's major systems stop synchronising as one organism.
Before Layer 4, the body can hide instability. It compensates, reroutes, buffers, and pushes through. But eventually the communication between systems begins to fracture. The nervous system, immune system, endocrine system, mitochondria, circadian rhythm, gut signalling, ECS tone, and behavioural state are still active — but they are no longer working together cleanly.
Neural rhythm, immune timing, endocrine pulses, mitochondrial membrane potential, circadian authority, vagal regulation, gut-brain signalling, ECS field timing, and behavioural state coherence.
Visible multi-system symptoms: sleep disruption, brain fog, pain sensitivity, inflammatory waves, autonomic instability, mood volatility, hormonal irregularity, and inconsistent energy.
Layer 4 is the unmasking layer. The problem did not start suddenly; the body simply ran out of compensation.
Systems Layer 5
Advanced Attractor Dynamics · ∆ Oracle
Simple metaphor
A storm that has lasted so long it creates its own weather system.
Layer 5 presents the theoretical idea that persistent dysfunction may stabilise into difficult-to-exit biological attractor states.
At this stage, QTOS explores whether some persistent biological states may become self-reinforcing through timing, signalling, inflammatory, metabolic, and behavioural feedback loops. In this interpretation, the persistence pattern itself may become part of the system dynamics.
Pattern flexibility, recovery reachability, system-level decision logic, relapse boundaries, attractor stability, and the ability to exit persistent dysfunction loops.
Relapse loops, repeating patterns, paradoxical responses, persistent dysfunction, and a sense that the system keeps returning to the same state despite surface-level correction.
Layer 5 is introduced publicly only as an advanced attractor-state hypothesis. Deeper theoretical material belongs in the protected book and reviewed-access material.
The Enzyme Systems Bridge
Physics-first biology describes biological systems as dynamic state spaces shaped by charge, coherence, rhythm, geometry, signalling, and adaptive reachability. Enzyme intelligence gives that framework a mechanism surface.
BioAtlas maps 1,724 enzymes across 38 families into six systems-state dimensions: signalling, oscillatory coordination, redox-regulatory, geometric/morphogenic, and informational/regulatory. This lets BioAtlas connect high-level systems-state ideas to enzyme-level biology: catalysis, signalling, tissue localisation, inflammatory context, epigenetics, ADME/BBB behaviour, ECS modulation, miRNA, synergies, evidence provenance, and graph-ready relationship intelligence.
Receptors, membranes, inflammatory tone, ECS modulation, tissue barriers, and electrochemical state.
Mitochondria, ROS, heme/flavin/NAD systems, oxidative pressure, and cofactor-linked biology.
Hormones, feedback, inflammatory pulses, ECS tone, cell-cycle timing, and pathway rhythm.
Electron transfer, proton movement, catalytic precision, conformation, and cofactor dependence.
Tissue architecture, matrix behaviour, stemness, regeneration, repair, and cancer morphology.
Epigenetics, miRNA, signalling logic, gene expression, pathway routing, and biological memory.
Boundary
The six enzyme field layers do not replace the Physics-First stack. They translate it into enzyme-level mechanism intelligence. This is a physics-informed systems annotation model, not a clinical diagnostic claim.
White paper DOI
This is the public DOI route for readers who want to view or download the physics-first disease dynamics paper directly from Zenodo.
Read on ZenodoQTOS DOI
The QTOS Vortex Architecture introduces the deeper framework: biology as coherence, disease as layered drift, and BioAtlas as the public platform layer built above the QTOS undercarriage.
Read on ZenodoThe stack beneath BioAtlas
The public page tells the story. The papers define the theory, architecture, guardrails, and computation logic. Reviewed access protects the deeper manuscript, diagrams, datasets, and diligence material.
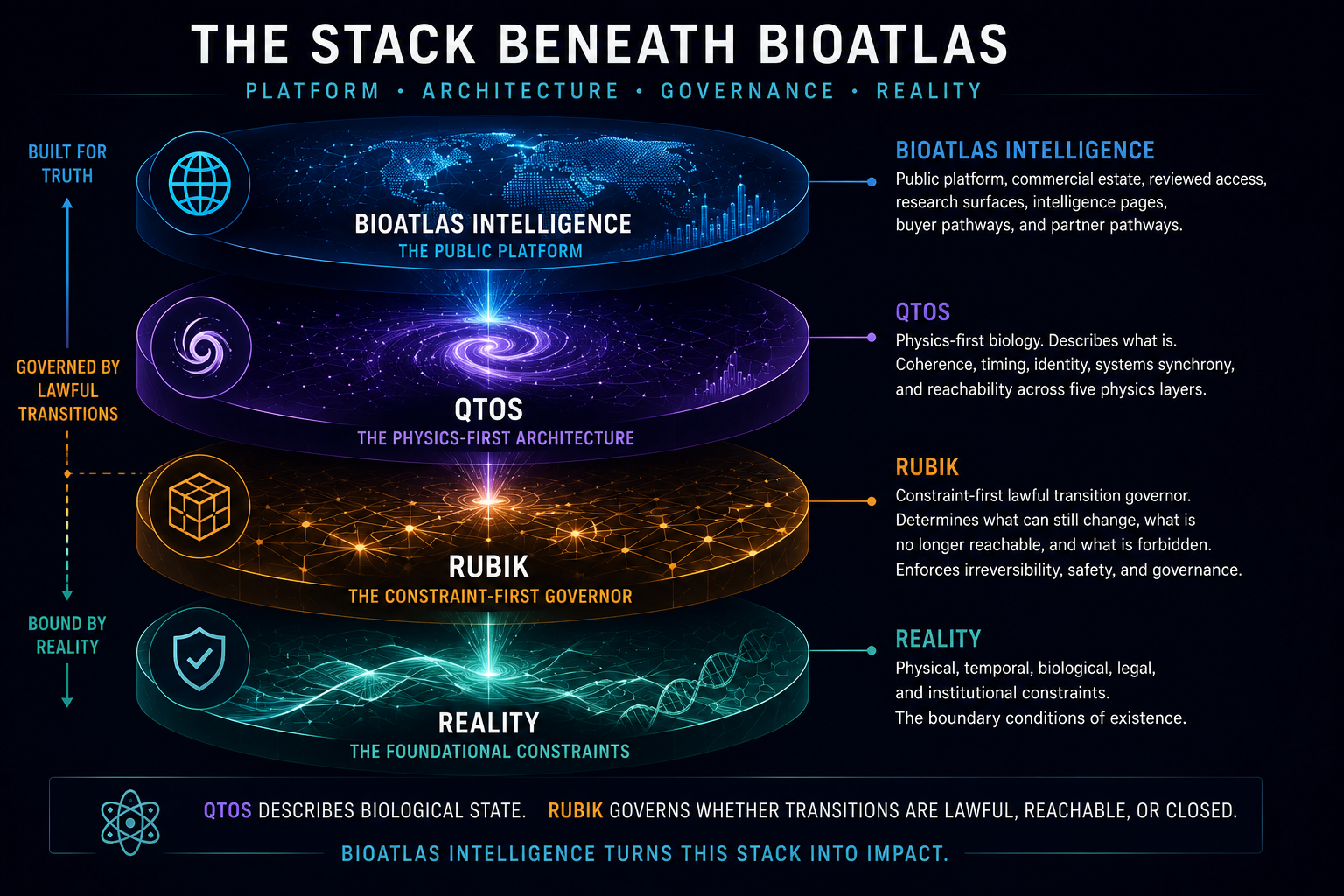
Public platform, commercial estate, reviewed access, research surfaces, and buyer pathways.
Physics-first biology, disease dynamics, coherence, timing, identity, and reachability.
Constraint-first lawful transition governance for irreversible systems.
Physical, temporal, biological, legal, and institutional constraints.
Public papers & technical notes
These public papers are routed through Zenodo so reads and downloads remain visible while deeper book visuals, datasets, intervention matrices, and diligence materials remain protected.
The biological theory paper
Introduces the physics-first interpretation of disease dynamics: coherence loss, timing drift, identity instability, system-wide desynchronisation, and advanced attractor behaviour.
Read on ZenodoThe core QTOS architecture paper
Defines the deeper QTOS architecture beneath BioAtlas: biology as coherence, disease as layered drift, and recovery as constrained state transition rather than simple reversal.
Read on ZenodoThe safety, scope, and non-claims paper
Defines the formal limits of QTOS: infrastructure, not diagnosis; constraint visibility, not prescription; descriptive governance, not autonomous intervention.
Read on ZenodoThe constraint-first computation paper
Positions Rubik beneath QTOS as the lawful transition governor for irreversible systems where recovery windows can close and brute-force computation may preserve the illusion of choice.
Read on ZenodoBelow QTOS: Rubik
QTOS describes biological state. Rubik sits beneath it as the constraint-first transition governor: the layer that asks whether a transition is still possible, no longer reachable, or structurally forbidden.
In irreversible systems, more computation does not restore choice once recovery windows close. Rubik is positioned as non-directive governance infrastructure for preserving truth under constraint, not as an AI, recommender, optimiser, or prediction engine.

Relevant public papers
These records form the public skeleton of the physics-first thesis: layered systems-state dynamics, QTOS, Rubik, constraint systems, irreversibility, signalling coordination, and exploratory systems-biology concepts.
Introduces the physics-first disease-dynamics framework across layered collapse, coherence, constraints, and state transitions.
Defines the QTOS capability envelope, limits, guardrails, non-claims, and infrastructure-grade scope.
Defines Rubik as a constraint-first computation and governance layer below QTOS, focused on reachability, impossibility, and deterministic boundaries.
Distinguishes descriptive mapping from constraint systems that define what is reachable, blocked, or structurally impossible.
Frames irreversibility, intervention limits, and biological state governance as a safety and systems problem.
Frames biophotonic regulation as an exploratory biological signalling layer within systems-oriented interpretation.
Frames DNA as a bioelectromagnetic interface for cellular intelligence and information-state interpretation.
Connects proton tunnelling, base-pair instability, epigenetic collapse, and oncogenesis as a systems biology research thread.
These records are included for provenance, publication, and review context only. They do not create medical advice, treatment instruction, dosing guidance, autonomous decision-making, or clinical access.
Forthcoming book
The public page should create desire without exposing the protected operating logic. The full QTOS manuscript expands each physics layer into diagrams, collapse maps, glyph operators, and deeper visual biology.
⊙
Boundary coherence, shielding, damping, and early field stability.
∞
Biological timing, oscillator synchrony, redox rhythm, and metabolic sequence.
Ψ
Identity flexibility, epigenetic rhythm, chromatin state, and miRNA timing.
✧
Signal-to-noise purification across whole-system biological rhythm.
∆
Advanced systems-state pattern interpretation and attractor analysis.
✪
Pathfinding through biological state-space and transition routes.
✷
Catalytic phase-change logic and disruption of rigid loops.
⟁
Large-scale systems interaction across interpersonal, behavioural, environmental, and organisational domains.
Visual gallery
The local image folder can be curated into a small public teaser set. The page should show only selected visuals from the forthcoming QTOS Physics-First Biology manuscript, while the full image library remains protected for the book release and reviewed access.
A public-facing summary of systems-state drift across timing, coordination, signalling, and adaptive biological stability.

A public-safe visual preview of Layer 1 systems-state instability and boundary coordination drift.

The ECS is presented as a gatekeeper layer because stress regulation, inflammatory coordination, and recovery flexibility affect biological reachability.
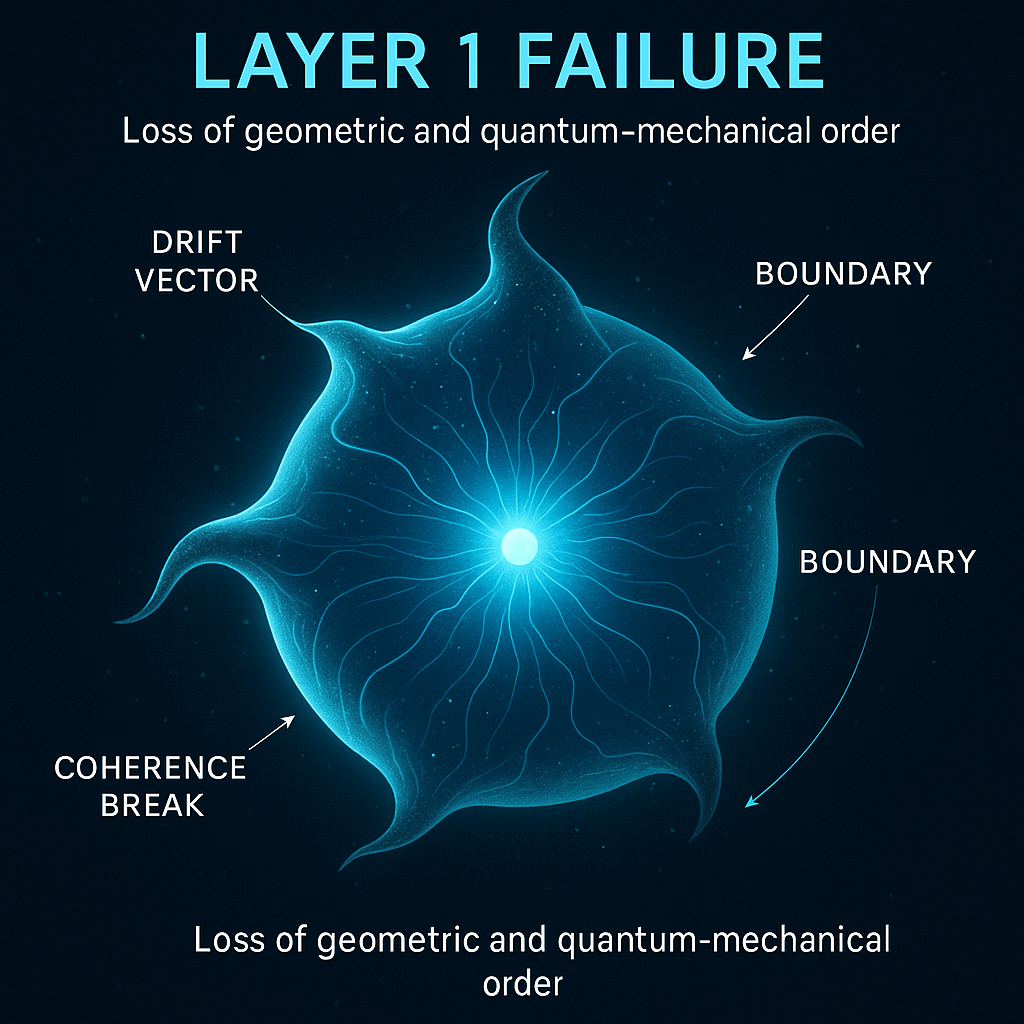
Layer 1 is framed as geometry-first collapse: boundary distortion, drift vectors, and coherence break before visible pathology.
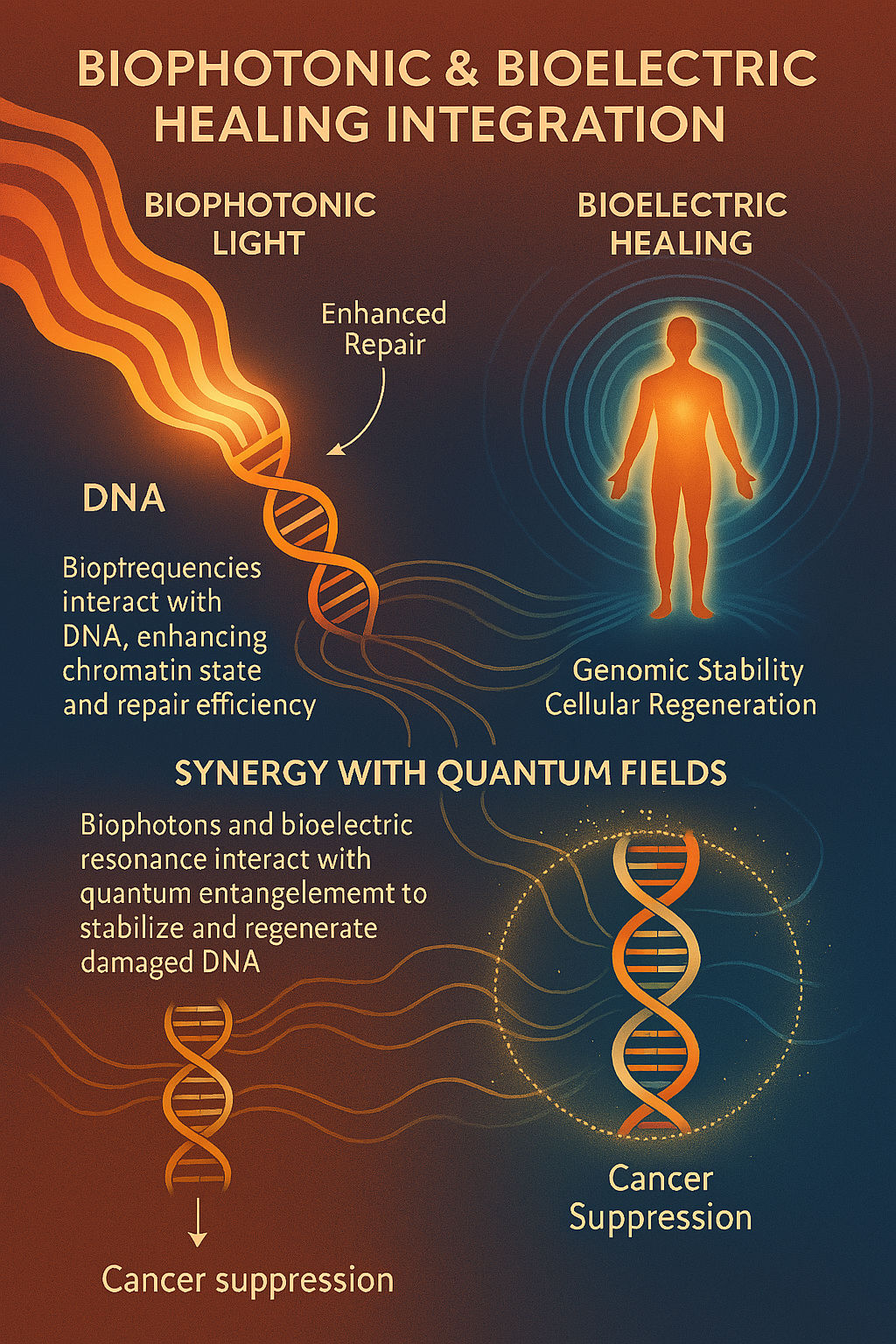
A public teaser of signalling-coordination themes explored within the QTOS manuscript.
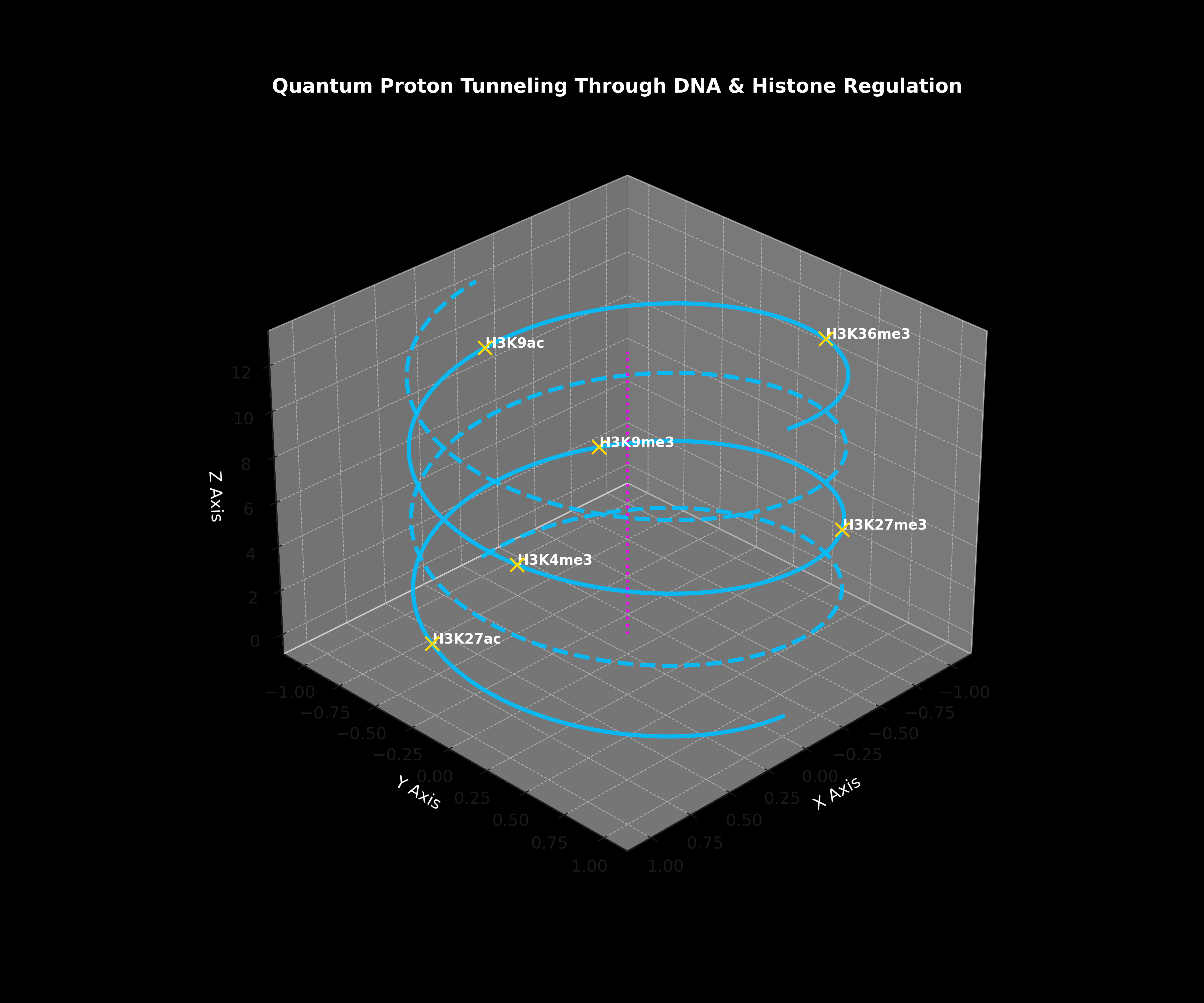
QTOS references proton-tunnelling concepts as part of its broader exploratory systems-state language.
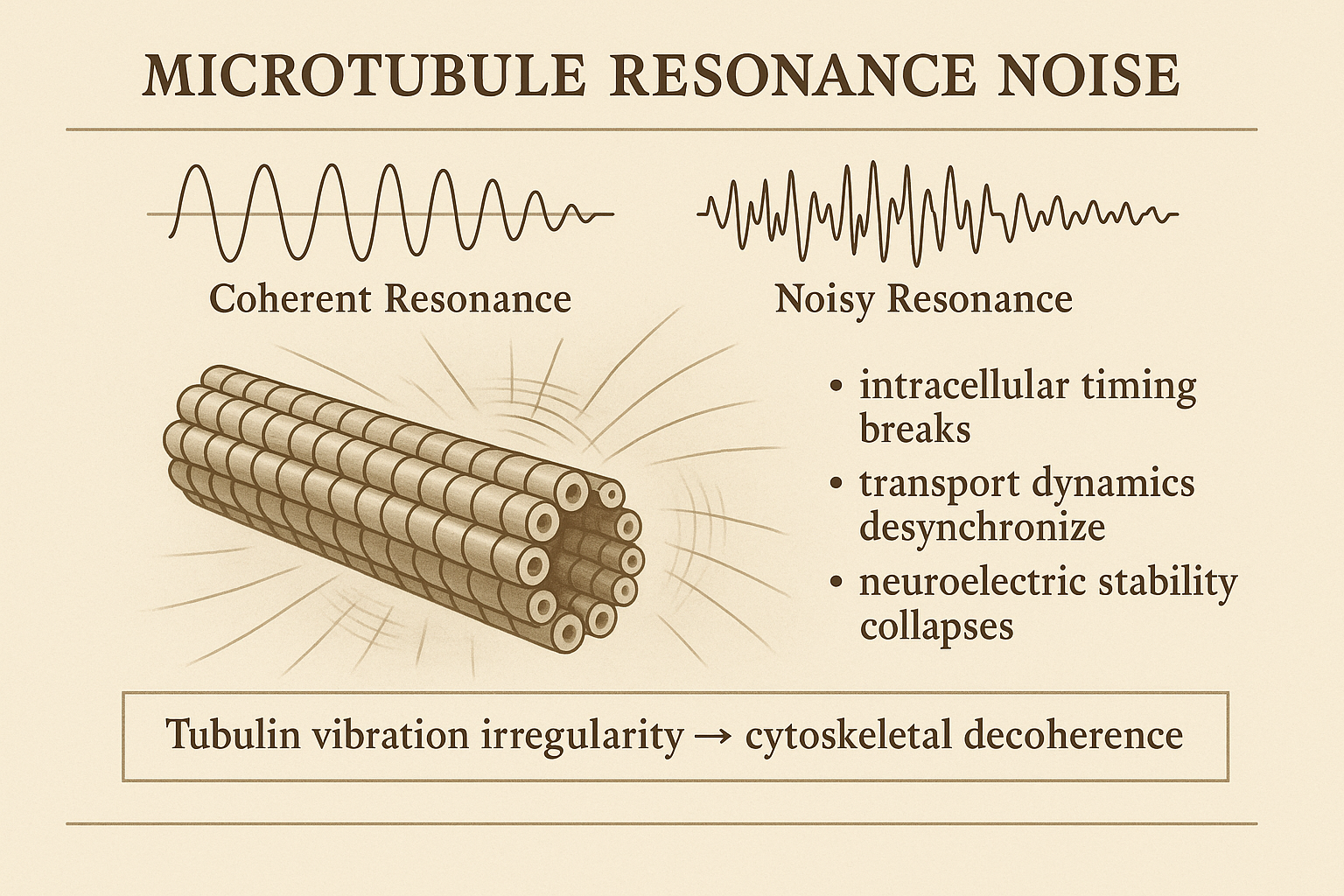
Microtubule resonance is used as a visual bridge between intracellular timing, structural organisation, and early instability.

A public-safe research and platform overview of the QTOS physics-first framework beneath BioAtlas Intelligence.
It is not medical advice, diagnosis, treatment instruction, clinical protocol, or a replacement for regulated clinical care.
Detailed intervention matrices, full manuscript diagrams, raw datasets, internal maps, book-only logic, and diligence material remain behind reviewed access. The capability-envelope paper defines the public guardrails, non-claims, and safety boundary.
Reviewed access
Researchers, investors, reviewers, collaborators, and strategic partners can request reviewed access. Public pages remain intentionally limited to protect the deeper book visuals, diagrams, datasets, and diligence-only materials. Public white papers are routed through Zenodo so reads and downloads remain trackable.